JARS v55n4 - What Is An Azalea?
What Is An Azalea?
Donald H. Voss
Vienna, Virginia
Reprinted from The Azalean, Vol. 20, No. 2, June 1998
At the 1997 Azalea Society of America convention, the editor of The Azalean suggested that an answer to the question, "What is an azalea?" would interest the membership. Firstly, the reader will not be surprised to see that azaleas are members of the plant kingdom ( Plantae ). They are seed plants in the division for flowering plants, the seeds of which are usually in closed carpels ( Angiospermae , or Magnoliophyta in newer terminology). When azalea seeds germinate, they produce two cotyledons ("seed leaves"), placing them in the familiar "dicot" class ( Dicotyledonae , or Magnoliopsida ). To approach the genus that includes azaleas ( Rhododendron ), we pass through the subclass Dilleniidae , the order Ericales , and the heath family ( Ericaceae ).
But what is an azalea? It can be a shrub or small tree. It may be evergreen or deciduous, and the inflorescences may be single- or multi-flowered. Typically, the evergreen azaleas are much branched shrubs, as broad as high, and clothed with medium- to dark-green, somewhat glossy, entire (not serrate or lobed) leaves. In winter, the foliage of some varieties takes on a bronzy or deep purplish coloration. While form and foliage are attractive year around, popular appreciation focuses on the often spectacular floral display in the spring. In the cultivated azaleas, flower form ranges from single to fully double, and color patterns may be solid, flecked, striped, sectored, or picotee. Colors included white, yellowish pinks, reds, pinks, purples, and purplish pinks. The deciduous azaleas are generally taller, upright plants with less branching than in the evergreen species. And, of course, in winter they present only the pattern of their stems and branches. The leaves are often much larger than those of the evergreen species, and they are usually not glossy. The flowers of most species have relatively long, narrow tubes. Colors include white and a variety of tones, from pale to strong, in the yellows, oranges, yellowish pinks, reds, pinks, purples, and purplish pinks.
That said, we must recognize the diverse morphological characteristics encountered in the various species of azalea. The flowers may be rotate-campanulate, campanulate, funnel-campanulate, funnel-shaped, or tubular-campanulate, and the number of stamens is usually five to ten. In the wild, the heights of familiar species of the evergreen azaleas range from prostrate to 3 m. (10 ft.) in height; their leaf-blade lengths, from 0.3 cm. (⅛ in.) to 8 cm. (3⅛ in.); and corolla length (receptacle to petal tip) from 0.6 cm. (¼ in.) to 6 cm. (2⅜ in.) Flower buds commonly produce one to four flowers, depending on the species.
In the deciduous azaleas, heights range from about 1 m. (3¼ ft.) to 10 m. (33 ft.); and leaf-blade lengths, from 3.4 cm. (1⅜ in.) to 12 cm. (4¾ in.). A flower bud may produce as few as three or up to twenty-four flowers, depending on the species. There are, of course, many other differences among the species in both the evergreen and deciduous groups. Rhododendron prinophyllum , for example, has a delightful clove scent. In short, there is no simple answer to the original question in terms of descriptive characters. But the pleasure that azaleas give as individual plants or integrated into a landscape is clear to those who know them.
The species and cultivars (cultivated varieties) of azalea, though widely adaptable, do exhibit certain differences in their cultural requirements. Cold hardiness, heat tolerance, the composition and pH of soil or other growing medium must be considered. Galle (1987) states that azaleas do best when the pH of the medium is between 4.5 and 6.0 (acidic). As with other groups of plants, a wide range of growth rates is encountered, partly reflecting the genetic makeup of the plants, and partly depending on climate, soils, availability of water, etc.
A Subset of Genus Rhododendron
The following notes indicate how azaleas came to be included in the genus
Rhododendron
, show
their relation to other parts of the genus, and delineate certain characteristics of the major groups
of azalea species. Azalea appeared among the genera embraced by Linnaeus (1753) in the
Species
Plantarum
(the starting point for modern botanical nomenclature), and - even after 1834, when
English botanist George Don incorporated azaleas in
Rhododendron
- the genus
Azalea
was maintained by a few botanists into the mid-1800s for evergreen azaleas and into the early 1900s
for deciduous azaleas.
Rhododendron
is one of 103 genera in the
Ericaceae
(heath
family), the general character of which is indicated in Table 1.
|
Table 1. Subfamilies of the
Ericaceae
(heath family), with illustrative examples of well-known
horticultural plants.
Subfamily Rhododendroideae Rhododendron (over 800 species including about 120 "azaleas") Epigaea repens (mayflower, trailing arbutus) Kalmia latifolia (mountain laurel) Loiseleuria procumbens (alpine azalea) Phyllodoce breweri (mountain heather) Subfamily Ericoideae Calluna vulgaris (Scotch heather) Erica cinerea (Scotch heath) Subfamily Vaccinioideae Andromeda polifolia (common bog rosemary) Arbutus unedo (strawberry tree) Arctostaphylos uva-ursi (bearberry, kinnikinik) Enkianthus campanulatus Gaultheria procumbens (wintergreen, checkerbox) Gaylussacia baccata (black huckleberry) Leucothoe fontanesiana (dog's hobble) Lyonia lucida (fetterbush) Oxydendrum arboreum (sorrel tree) Pieris japonica (Japanese andromeda) Vaccinium corymbosum (high-bush blueberry) |
A brief history of Rhododendron classification was published by the Philipsons in 1974. The road from Species Plantarum to the modern classification of the genus (which includes all of the azaleas) is long and exceedingly tortuous. We will not travel that road but note here several factors to consider when using the modern classification:
The Linnean system was based primarily on a numerical, "sexual" classification reflecting the number, length, and union (if any) of the stamens and the number of styles in each flower. These criteria made it easy to sort plants into the various "boxes" of the classification system. But when this system is used and the contents of a particular box are examined, the plants are often disparate with respect to characters other than the number and type of the sexual parts.
Linnaeus' contribution of binomial nomenclature (genus name + specific epithet, instead of the unwieldy phrase names previously used) was widely adopted and is still in use today. Thus the Linnean name Azalea indica replaced names for that plant such as Breyne's "Chamaerhododendron exoticum, amplissimis floribus liliaceis." Linnaeus' artificial sexual system of classification was, however, soon challenged. Systems based on "natural" relationships reflecting morphological affinities emerged. De Candolle, for example, divided the dicotyledonous plants into those with and without a corolla, and those with a corolla into groups with separate petals and with united petals.
In England, meanwhile, G. Don incorporated consideration of morphological characters into his classifications and moved azaleas into genus Rhododendron . He divided the genus into sections, based largely on geographical origin for the azaleas. After Darwin's concepts of natural selection spread in the latter half of the 19th century, botanists increasingly focused on evolutionary development as a basis for classification. In the same period, large numbers of species new to Western science were being introduced as plant exploration expanded in the Himalayan region, China, Japan, and elsewhere in Asia.
Changes in classification of Rhododendron (including the azaleas) have continued to the present. One scheme familiar to rhododendron enthusiasts is the grouping of many species that exhibit certain similarities into "series," a scheme promulgated by Prof. Isaac Bayley Balfour at Edinburgh circa 1920. A treatment integrating the Balfour system and other developments in rhododendron classification appeared in a 1949 article by the German botanist Sleumer. As subsequently modified by Sleumer and other, this treatment remains central to taxonomic work in the genus.
The most comprehensive modern taxonomic and nomenclatural treatment of the rhododendrons and azaleas is the "Edinburgh Revision" (see Reference 4).
In 1753, Linnaeus listed six species in his genus Azalea: indica (India [Orient]), pontica (Ponto, Trapezunte [NE Turkey, NE Asia Minor]), lutea (Virginia [east-central North America]), viscosa (Virginia), lapponica (alpibus Lapponica [Lapland mountains]), and procumbens (alpibus Europae [European alps]). These were placed in his category "Pentandria [five stamens] Monogynia [one style]," which includes plants with perfect flowers that have stamens not joined together and not determinately proportioned in length. In contrast, the five Linnean species of Rhododendron were placed in "Decandria [ten stamens] Monogynia." The six Linnean species of Azalea are now classified as follows:
| Linnean Name | Current Name |
| Azalea indica L. | Rhododendron indicum (L.) Sweet |
| A. pontica L. | R. luteum Sweet |
| A. lutea L. | R. periclymenoides (Michaux) Shinners |
| A. viscosa L. | R. viscosum (L.) Torrey |
| A. lapponica L. | R. lapponicum (L.) Wahlenberg |
| A. procumbens L. | Loiseleuria procumbens (L.) Desvaux |
Rhododendron indicum is the familiar evergreen azalea; R. luteum , R. periclymenoides , and R. viscosum are deciduous azaleas. R. lapponicum is a lepidote (scaly) rhododendron. Finally, the Linnean A. procumbens is now included in genus Loiseleuria .
We remind the reader that G. Don transferred azaleas into genus Rhododendron in 1834, but we will not trace the many subsequent changes in azalea nomenclature. Among the species in the azalea complex (azaleas in the broad sense), are many that most of us have never heard of nor seen. To a considerable extent, this reflects a large increase in the number of Asian species, particularly Chinese, in recent decades.
| Table 2. Provenance of evergreen azaleas as shown in the Edinburgh Revision. | ||||||
| Subgenus | No. species | China | Japan | Other Asia | North America | Europe/Near East |
| Azaleastrum | 16 | 14 | 2 | 6 | -- | -- |
| Mumeazalea | 1 | -- | 1 | -- | -- | -- |
| Candidastrum | 1 | -- | -- | -- | 1 | -- |
| Therorhodion | 2 | 1 | 1 | 2 | 1 | -- |
| Tsutsusi | 80 | 53 | 27 | 7 | -- | -- |
| Pentanthera | 22 | 1 | 5 | 1 | 15 | 1 |
Provenance
The geographical origin of the species comprising the azalea complex in the subgenera
of
Rhododendron
makes clear the predominance (in terms of number of species)
of China (including Taiwan and Hong Kong) for the evergreen species and North America
for the deciduous. If we consider only the evergreen species most familiar in horticulture,
Japan is the dominant source. The summary (Table 2) is based on the provenance of
these species as shown in the Edinburgh Revision. The geographic-distribution numbers
may add to more than the number of species, because a given species may be native in
more than one area (for example,
R. simsii
is native to Upper Burma, China, Laos,
Thailand, and Japan's Ryukyu Islands). Because many of the new Chinese species are unfamiliar
even to western botanists specializing in the genus, it is possible that some may later be
combined when more specimens become available for study. And, of course, additional species
may be found.
Vestiture
An interesting scheme for distinguishing groups within genus
Rhododendron
is based
on the nature of the plants' vestiture (scales, glands, and/or hairs). With the aid of a
hand lens (at least 10x power), one can usually place a plant from the genus into one
of three major groups. From detailed study of about one-third of the species in
Rhododendron
, German botanist Almut Seithe, as reported in the proceedings of the
1978 International Rhododendron Conference, identified forty-three types of vestiture in
the genus. These can be subsumed in four major categories: virgate (straight, wand-like)
hairs; flock (branched) hairs; glands; and scales. The hairs in these categories may be
identified as unicellular or multicellular, as well as by other variations. The categories
of vestiture relate to the major groups in Rhododendron proposed by Seithe (at a new rank -
"chorus" - between genus and subgenus) in the following manner (see also Figure 1, reproduced
with the kind permission of Dr. Seithe):
-Chorus subgenerum Rhododendron (scaly rhododendrons) has scales and virgate hairs
-Chorus subgenerum Hymenanthes (non-scaly rhododendrons) has flock hairs and glands
-Chorus subgenerum Nomazalea (the azalea complex) has virgate hairs and glands
This does not mean that each member of a chorus subgenerum has both types of vestiture specified. Thus the evergreen azaleas generally have multicellular hairs, often flattened and adpressed; some species (e.g., R. oldhamii Maxim. and R. stenopetalum (Hogg) Mabb. [syn R. macrosepalum Maxim.) also have stalked glands. In the deciduous azaleas, both unicellular and multicellular hairs are found, the latter often gland-tipped.
Sleumer did not accept the evolutionary implications of the chori subgenerum and noted exceptions to Seithe's categories. Chamberlain (Edinburgh Revision, Ref. 4B) noted that stiff, unbranched multicellular hairs may be found in five of the twenty-four subsections in Hymenanthes . Nonetheless, Seithe's groups provide a useful, if tentative, guide for many of the species encountered in horticulture. Using this method shows, for example, that Rhododendron mucronulatum Turcz. is a lepidote rhododendron, not an azalea.
The detailed nature and location of vestiture may be important in the identification of a species. This is particularly true for the deciduous azaleas, where gland-tipped hairs are often diagnostic. Even if one is not concerned with the details of vestiture as an aid to identification, examination of shoots, buds, leaves, and flowers with a lens reveals an aspect of beauty often not appreciated in these plants.
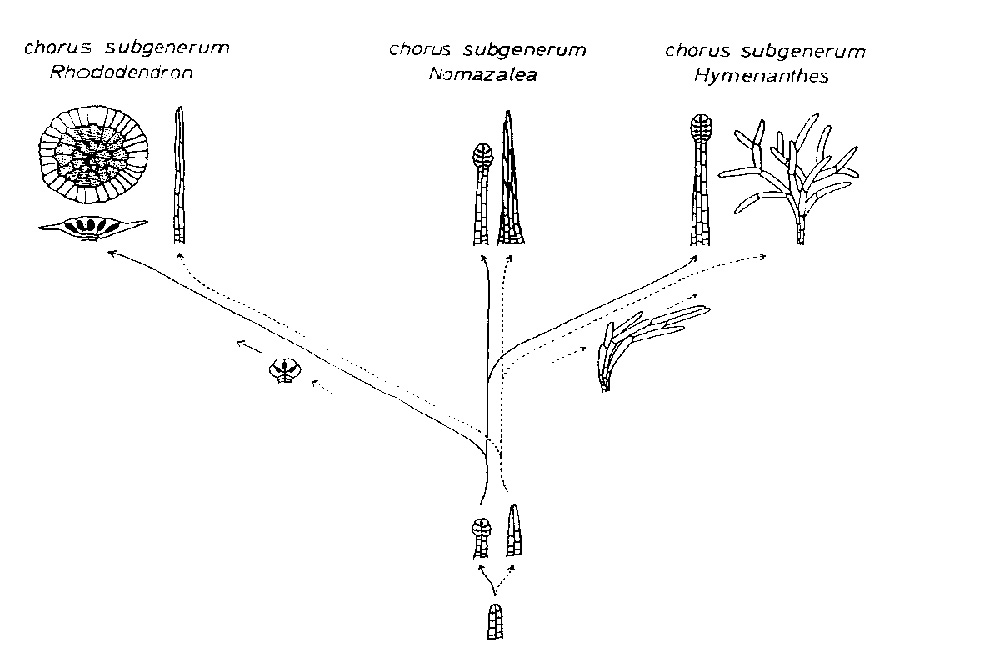
|
|
Figure 1. Phylogenetic Tree of the Chori subgenerum and the hair class variants.
chorus subgenerum Rhododendron with scales and virgate hairs chorus subgenerum Nomazalea with glands and virgate hairs chorus subgenerum Hymenanthes with glands and flock hairs Reprinted with permission from Contributions Toward a Classification of Rhododendron, page 115 (Figure 13), copyright 1980, The New York Botanical Garden. |
Current Rhododendron Classification
The subgenera and their subordinate sections in genus
Rhododendron
are shown in Table 3.
For each of these taxa, the "type species" is identified, and the number of included species
is listed. In some cases, examples of included species are shown and information is given on
the forms of inflorescence encountered.
|
Table 3. The subgenera of Rhododendron L.
Note: The number of species (spp.) noted for each subgenus and section are from "The Current Classification" section of the introduction to Chamberlain, 1996. If validly published hybrid species and certain "accepted names" are included, the count in major categories would be about 50 percent greater. Plants commonly called "rhododendrons" [Scaly rhododendrons] Subgenus Rhododendron [Type: R. ferrugineum L.]; ca 360 spp.
Inflorescence always a raceme, most frequently
several-flowered, with very short rachis (umbellate), sometimes reduced to a single flower, or with
elongated rachis (nonumbellate)
Incl.
R. dauricum
,
R. ferrugineum
(alpine rose),
R. hippophaeoides
,
R. impeditum
,
R. keiskei
,
R. micranthum
,
R. mucronulatum
,
R. myrtifolium
,
R. racemosum
Inflorescence always a terminal raceme, few- to many-flowered,
lax or dense; often forming a more or less spherical "truss" of flowers
Inflorescence lateral below terminal or pseudoterminal
vegetative buds, with 1 to several flowers
Inflorescence lateral, below vegetative buds, with 1 to 2 flowers
Inflorescence lateral, below vegetative buds, with 1 to 2 flowers
Inflorescence buds terminal, flower stalks bearing leaf-like
bracts, with 1 to 3 flowers
Inflorescence and vegetative buds enclosed in same terminal bud scales
Incl. R. eriocarpum , R. indicum , R. kaempferi , R. kiusianum , R. nakaharae , R oldhamii , R. ( ripense x stenopetalum ) 'Mucronatum', R. simsii , R. stenopetalum (syn. R. macrosepalum ), R. tashiroi , R. yedoense
Flowers solitary or in subumbellate racemes with up to 15 flowers,
flowering time varies
Incl.
R. amagianum
,
R. dilatatum
Flowers solitary or in subumbellate racemes with up to 4 flowers, flowering
before or with leaf expansion
Flowers terminal on previous-year branchlets; flowers before, with, or
after expansion of leaves; vegetative shoots below terminal buds, or in some cases from lowest scaly leaves of
terminal bud ["scaly" here does not mean lepidote]
Inflorescence a shortened terminal raceme, flowers before, with, or after leaf expansion Subsection Sinensia (Nakai) K. Kron [Type: R. molle (Blume) G. Don]; 1 sp. Incl. subsp. molle and subsp. japonicum (A. Gray) K. Kron
Subsection Pentanthera G. Don [Type: R. luteum Sweet];14 spp.
Incl.
R. luteum
from the Caucasus, Turkey, and Eastern Europe
and the North American
R. atlanticum
,
R. calendulaceum
,
R. periclymenoides
,
R. viscosum
, etc.
Inflorescence an umbellate terminal raceme of 1 to 6 flowers,
flowers before or with leaf expansion
Inflorescence an umbellate terminal raceme of 6 to 15 flowers,
flowers with or after leaves expand
Inflorescence an umbellate terminal raceme of 3 to 15 flowers, flowers before leaves expand |
Type species relate primarily to the technical requirements governing the creation of botanical names. Thus, a type species is associated with the name of each family, genus, or infrageneric taxon. While the type species is part of a subgenus or section, one must understand that it is not necessarily typical of those found in the subgenus or section.
In any named taxon (a group of plants such as a species), there will be a range of variation in morphological characters - leaf shape and size, calyx and corolla size and form, color, etc. In some cases, a type species may have characteristics closer to an extreme of the range rather than be representative of the average. That said, many type species provide useful exemplars for visualizing the plants in a subgenus or section with which they are associated.
The taxonomic classification of Rhododendron is based on above-ground morphological features. Below ground, members of the genus generally have compact, fibrous root systems - but there are exceptions. Some of the lepidote (scaly) species are epiphytes. In the deciduous azaleas, as Solymosy (1976) pointed out, several (notably including R. viscosum (L.) Torr.) have shallow roots with runners sometimes 10 m. (more than 30 ft.) long! These runners send up shoots that develop into new plants. Galle also discusses the often sparse, widespreading root systems of our native deciduous azaleas and gives valuable cultural advice for their transplantation.
The overlapping ranges of flower forms within the various subgenera and sections means that listing the forms occurring in each would not be useful in visualizing differences between these groups. In general, flower forms in Rhododendron range from rotate to campanulate or funnel form, sometimes tubular (or combinations of these forms), and the flowers are usually slightly irregular (zygomorphic, bilaterally symmetrical).
The nature of the inflorescences, in contrast, provides some important clues to taxonomic relationships. Three terms used in describing these inflorescences are raceme, rachis, and umbellate. A raceme is a simple inflorescence of stalked flowers on an axis supporting the flowers. This axis is called a rachis and may be relatively long or short. If the rachis is short, the flower stalks will appear to arise from nearly the same point, producing a compact inflorescence. This appearance is referred to as umbellate (resembling an umbel), because an umbel is an inflorescence in which the flower stalks arise from a common point. (Think of the primary branching of the Queen Anne's lace flower, a compound umbel.) Terminal buds are located at the end of branchlets; lateral buds are located in the leaf axils below the terminal buds.
At this point, readers who desire specific information about the various species and cultivars
of azaleas should refer to Galle's Azaleas. In that work, thousands of azaleas are described and
many are pictured in color. An alternative would be to stroll among the azaleas and reflect on
the words of Keats:
A thing of beauty is a joy forever.
Its loveliness increases; it will never
pass into nothingness
Endymion
References
1. Argent, George, et al. 1997.
The Rhododendron Handbook 1998: Rhododendron Species
in Cultivation
. London: The Royal Horticultural Society.
2. Chamberlain, David, et al. 1996.
The Genus Rhododendron; Its Classification & Synonymy
.
Edinburgh: Royal Botanic Garden Edinburgh.
3. Don, George. 1834.
General History of Dichlamydeous Plants
. London.
4. Edinburgh Revision of Rhododendron:
(A) Cullen, James. 1980. A Revision of Rhododendron:
I. Subgenus Rhododendron Sections Rhododendron & Pogonanthum.
Notes from the
Royal Botanic Garden Edinburgh
. 39(1):1-207.
(B) Chamberlain, D. F. 1982. A Revision of Rhododendron: II. Subgenus Hymenanthes.
Notes from the Royal Botanic Garden Edinburgh
. 39(2):209-486.
(C) Philipson, W. R. and M. N. Philipson. 1986. A Revision of Rhododendron: III: Subgenera
Azaleastrum, Mumeazalea, Candidastrum, and Therorhodion.
Edinburgh Journal of Botany
.
44(1):1-23.
(D) Chamberlain, D. F. and S. J. Rae. 1990. A Revision of Rhododendron; IV. Subgenus Tsutsusi.
Edinburgh Journal of Botany
. 47(2):89-200.
(E) Kron, K. A. 1993. A Revision of Rhododendron: V. [Subgenus Pentanthera] Section
Pentanthera.
Edinburgh Journal of Botany
. 50(3):249-364.
(F) Judd, W. S. and K. A. Kron. 1995. A Revision of Rhododendron: VI. Subgenus Pentanthera
(Sections Sciadorhodion, Rhodora, and Viscidula).
Edinburgh Journal of Botany
. 52(1):1-54.
5. Galle, Fred C. 1987.
Azaleas
. Rev. & enl. ed. Portland, OR: Timber Press.
6. Philipson, W. R. and M. N. Philipson. 1974. A History of Rhododendron Classification.
Notes from the Royal Botanic Garden Edinburgh
. 32:223-238.
7. Seithe, Almut. 1980. Rhododendron Hairs and Taxonomy. In Luteyn, James L., ed. 1980.
Contributions Toward a Classification of Rhododendron
. New York: New York Botanical Garden.
8. Sleumer, H. 1949. Ein System der Gattung Rhododendron L.
Bot. Jahrb
. 74:511-553.
9. Solymosy, Sigmond L. 1976. A treatise on native azaleas.
Bulletin of the Louisiana Society for
Horticultural Research
. 4(2):16-17.
1 The personal name(s) or abbreviated names following botanical names identify the author(s) of those names. For example, Azalea indica L. means that Linnaeus (abbreviated "L.") applied that botanical name to the species. When the specific epithet indica was transferred to the genus Rhododendron by Sweet, "(L.)" was included after the new combination to show that the epithet was originally applied by Linnaeus. The gender of the epithet was changed to match that of the genus, hence Rhododendron indicum .
Don Voss, a member of the Potomac Valley Chapter, is a frequent contributor to the Journal on Rhododendron nomenclature. He is a volunteer at the herbarium of the U.S. National Arboretum.
How Many Stamens?
In discussing "What is a 'single' flower? with the author, William C. Miller III raised an interesting issue.
If the "normal" azalea flower is said to have five each of sepals, corolla lobes, and stamens, is a flower
departing from this model by having additional stamens (e.g., seven) therefore not single? With respect to
the evergreen azaleas in
Rhododendron
subgenus
Tsutsusi
, the answer is "no."
In section Tsutsusi of the subgenus, two-thirds of the 65 species (41) have a normal complement of five stamens: 14 usually have 10 stamens. For the remaining species, the normal count varies in ranges such as 4-5, 4-6, 6-10, 7-10, 8-9, 9-10, etc. And these are the species! Mix the genes in a stew-pot of interspecific hybridity and one should not be surprised by stamen counts other than five. To illustrate the variability, the usual stamen count for several species familiar in horticulture is tabulated below:
indicum 5
nakaharae 10
kaempferi 5(-6)
oldhamii (8-)10
kiusianum 5
simsii (8-)10
stenopetalum 5(-7)
yedoense 10
ripense 10
Note: The Rhododendron Handbook 1998 accepts R. stenopetalum (Hogg) Mabb. as the name of the species more commonly known as R. macrosepalum Maxim. 'Mucronatum' is now considered to be R. ( ripense x stenopetalum ). The bottom line: in distinguishing flowers as "single" or "double," concentrate on the condition of the stamens (normal, with functional anthers; partly petaloid; or full petaloid), and - especially in hybrids - accept as normal some variability in number of stamens. Similarly, while the normal complement of corolla lobes is five, a plant that produces a corolla with six or even seven lobes may fall in the range of variability encountered in "single" azalea flowers.